(to read part 1 of this series click here, to read part 2 of this series click here)
Modern Humans
Modern humans near-universally live with permanently compromised fascial systems.
The lone exceptions– isolated hunter-gatherer tribes– may point to the root of this disorder, in the shift away from the lifestyle to which humans evolved. A 2015 study of the Hadza people– a group of hunter-gatherers living in north-central Tanzania– highlights this loss of biomechanical efficiency. This study tracked 30 adult Hadza population members via GPS and breath monitors (to follow location/ metabolism), while the study members had their urine tested regularly to see how quickly they passed chemically altered water (to further test metabolism/ energy usage). The study found that, despite a far more active lifestyle (roughly 50% more distance walked than average westerners, plus vigorous exercise in hunting/ gathering), the Hazda people burned no more calories than equivalently-built western individuals. In other words, a Hadza person could travel 50% further (with far greater overall activity) than a same-sized westerner, while burning the exact same number of calories. This is the most literal definition of efficiency.
This phenomenon is not just contained to humans. Mammals living in domesticity- whether its zoo animals, pets, or farm animals– show similarly chronic inefficiency. According to the author of a recent study on primate energy levels, “comparisons of wild and captive primate populations indicate similar levels of energy expenditure”, just like in the above Hadza study.
In other words, captivity (or domesticity) correlates with decreased activity output from the same energy input. Biomechanical efficiency appears to decrease by the shift away from traditional lifestyles towards sedentary lifestyles. It therefore seems likely that modern humans became chronically inefficient in the shift from hunter-gatherer lifestyle to modern agricultural/ sedentary lifestyle.
But regardless of the source of the condition, modern humans live with fascial systems that have become adapted to chronic inefficiency.
Living with Fascial Inefficiency
Mammals evolved to move, as both an advantage and a necessity. Mammals are the most capable and versatile movers on the planet. However, as discussed in part 2 of this series, the complex movement capabilities of mammals are underscored by a system of densely organized wires running perpendicular to tendons (skin ligaments). Free unrestrained movement (stretching, running, grooming) keeps these wires unencumbered by constantly applying tension to each of the wires. But when movement becomes restrained– whether it’s a monkey in a cage, or a child sitting in class– inefficiency follows.
There is a feedback mechanism evolved into the capabilities of fascia– feedback which allows wild mammals to know when to stretch, where to groom– how to move to keep their fascia optimized and healthy. The fascial system is not only comprised of wires and levers (skin ligaments and muscle tissue), but is also innervated with proprioceptive nerve bundles. Proprioception is the ability of the muscle system to recognize itself, to self-analyze. Proprioception is how you can know where your arm is, even without looking. And proprioception becomes compromised as the efficiency of the fascial system lessens.
A monkey banging on bars, or a child fidgeting in their chair, is likely experiencing some level of proprioceptive distress– their fascial system is telling them to move, to free up encroaching entanglements. But whether it’s restriction via zoo bars or a strict teacher, their system is not allowed free unrestrained movement to self-maintain. So entanglements occur, which then lessen the fascial system’s ability to self-analyze in the future (by compromising blood-flow to proprioceptive nerve bundles). Inefficiency begets more inefficiency.
Blood and lymphatic vessels travel through fascia. So when entanglements occur, blood/ lymph flow is redirected. In an otherwise healthy fascial system, this is beneficial– bodily fluids are forced (by entanglements) to travel through areas of inefficiency, helping to rebuild areas that have become inefficient due to trauma. But when inefficiency is widespread and chronic, bloodflow is compromised on a system-wide level, lowering the ability of the system to direct blood to where it is needed on demand (whether due to activity or due to trauma). The ability of the whole system to adapt and self-maintain becomes chronically compromised.
Modern human fascial systems have adapted to these conditions by showing preferential treatment to certain areas over others. Imagine, for example, a marionette where the wires are tangled. If you were to consistently pull on some of the wires but not others, knots would form, tighten, and eventually the wires on the side that you consistently pull would allow you to control the movement of that side. However, every time you pulled those wires, other parts of the marionette would also move, due to the entangled connections to those other areas.
This is a gross simplification of the function of fascia in modern humans. Certain areas become relatively efficient– the areas where you most consistently pull the wires. These areas show the most linear connection between spinal action and limb action. But because of fascial entanglements and muscle knots, extra movement is generated in areas adjacent to the intended area– the areas where the wire has been shortened and borrowed against. This is what is meant by ‘biomechanical borrowing’– areas of efficiency are generated by borrowing wire from adjacent areas. The areas of greatest use gain efficiency by pulling wire from areas of least use.
Since chronic inefficiency begins during childhood, biomechanical development can become compromised. The areas of greatest use will have tendons that reach their full length, due to the extra (borrowed) give in the wire. But the areas of least use will stay chronically tight, restricting blood/ lymph flow and thereby becoming stunted over time.
In addition, areas of little use tend stay little used, due to lacking strength and dexterity (from shortened inefficient tendons). For example, how often do you use your dominant hand, vs your non-dominant hand? When an area feels capable/ strong it gets used more often, and weaker/ less capable areas are ignored. Efficiency begets efficiency and inefficiency begets inefficiency. So blood/ lymph are rarely directed to little-used areas, and when they are so directed, only travel very slowly (due to being routed via entanglements through areas of greater use). Tendons that don’t reach full length by around age twenty-five (when the biomechanical system fully matures) will remain relatively inefficient for life.
Analyzing Biomechanics Visually
A measure of biomechanical efficiency in an otherwise inefficient system is a measure of how cleanly spinal action translates to limb action. If the tendons are full length and the fascia remain slack when spinal action is initiated, the area is efficient– spinal action will translate fully to limb action, with the fascia loose and ready to supplement/ complement this action at will. If by default the tendons are shortened and the fascia are taut, the spinal action will be stymied/ redirected away from the intended limb action by fascia that are interceding involuntarily.
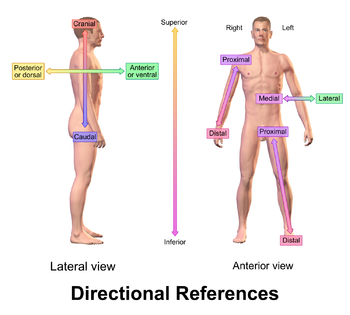
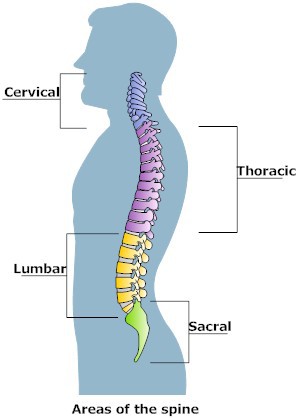
There are two main biomechanical pathways (medial and lateral) in each of the five spinal areas (cervical, thoracic, lumbar, sacral, and coccygeal), and each spinal area has separate anterior and posterior muscles (to read about anterior vs posterior efficiency, click here). The three main spinal areas for biomechanical study are cervical (the vertebrae/ muscles of the neck), thoracic (the vertebrae/ muscles of the chest/ abdomen/ upper/middle back), and lumbar (the vertebrae/ muscles of the lower abdomen/ lower back). These areas can be seen in the image below
In the previous example of the concert pianist from part 2, his thoracic areas were fully efficient– his thoracic spinal action transferred with full force to his hands, with fascia along the way loose unless directly engaged. He was fully efficient in both medial and lateral areas (thumb/ forefinger are medial, while the middle/ ring/ pinky fingers are lateral), and overall, he favored anterior development over posterior development (although both were fully efficient in his thoracic areas).
In total, there are twelve broad measures of biomechanical efficiency that are able to be identified visually– the efficiency of the cervical-attached areas (head/ neck), the thoracic-attached areas (chest/ back/ arms/ hands), and the lumbar-attached areas (lower back/ abdomen/ legs/ feet). These are subdivided into medial (close to the spine) and lateral (further from the spine) biomechanical pathways. And they originate as either anterior (front of the body) or posterior (back of the body) spinal action.